¢|Š®ę░▄į└“├ń─ŠĄ──═║«ąį蹊┐ |
| «öŪ░╬╗ų├: ųąć°├ń─ŠŠW > įį┼Ó╝╝ąg > |
| üĒį┤Ż║╬┤ų¬ ū„š▀Ż║admin ░l▓╝ĢrķgŻ║2013-07-28 07:21 |
|
¢|Š®ę░▄į└“Ż©Styrax tonkinensisŻ®Ż¼ėų├¹įĮ─Ž░▓ŽóŽŃĪó░ū╗©śõŻ¼ī┘░▓ŽóŽŃ┐Ų░▓ŽóŽŃī┘┬õ╚~å╠─ŠŻ¼ų„«aįĮ─Ž¢|Š®×│Ż¼į┌╬ęć°ų„ę¬Ęų▓╝ė┌įŲ─ŽĪó┘Fų▌ĪóÅV╬„ĪóÅV¢|Īó║■─ŽĪóĖŻĮ©║═ĮŁ╬„Ą╚Ąž[1 ~ 3]ĪŻį┌įŁ«aĄžų„ę¬╔·ķLį┌║Ż░╬100 ~ l 000 mĄ─Ą═╔ĮŪ┴ĻĄžÄ¦Ż¼Ž▓رØÖ╩Ķ╦╔Ę╩╬ų═┴╚└Ż¼ę▓─═žÜ±ż║═─═Ė╔įń[4 ~ 5]ĪŻįōśõĘN╦┘╔·Ż¼Š▀║▄ÅŖĄ─├╚č┐Ė³ą┬─▄┴”Ż¼śõĖ╔═©ų▒Ż¼─Š┘|░ū╔½╝Üų┬Ż¼▓─▌p▄øŻ¼─Š└wŠS▌^ķLŻ¼╩Ū└ĒŽļĄ─╣ż╦ćĪó─z║Ž░Õ║═╝łØ{ė├▓─Ż╗ŲõĘNūė║¼ėą╚╦¾w╦∙▒žąĶĄ─ė═╦ß║═üåė═╦߯¼Š▀ėą▌^Ė▀Ą─╩│ė├ārųĄŻ¼śõų¼║¼ŽŃų¼╦߯¼×ķ┘Fųž╦Ä▓─Ż¼╩Ūę╗ĘN║▄ėą░lš╣Ū░═ŠĄ─╩│╦Äė├ė═┴ŽśõĘN[3 ~ 4]Ż╗Ųõ╗©░ū╔½│╔┤«Ż¼ŪÕŽŃŻ¼ę▓╩Ūę╗ĘNā×┴╝Ą─ŠG╗»śõĘN[6]ĪŻ 999├ń─ŠŠW,999miaomu.com 1 ▓─┴Ž┼cĘĮĘ© 1.1╣®įć▓─┴Ž╝░Ųõ╠Ä└Ē ╣®įć▓─┴Ž×ķĘNį┤▓╔ūįĮŁ╬„╦ņ┤©Ż¼į┌ĮŁ╠K╩ĪµéĮŁ╩ąōPūėĮŁ┴ųśI┐Ų╝╝ėąŽ▐╣½╦Š├ńŲį╔·ķLĄ─¢|Š®ę░▄į└“ę╗─Ļ╔·▓źĘN├ńų”ŚlĪŻ īó¢|Š®ę░▄į└“ų”Śl╚¶Ė╔Ęųäeį┌-7ĪµĪó-18Īµ║═-25ĪµŽ┬╠Ä└Ē, Ģrķg×ķ 24Īó 48Īó 72 hĪŻęį5Īµ×ķī”šš, ╠Ä└ĒĮY╩°║¾Ęųäe╚Īų”ŚlĄ─ĒgŲż▓┐Ż©▒▄ķ_č┐č█Ż®▀Mąą£yČ©ĪŻ 1.2įć“×ĘĮĘ© ┐╔╚▄ąį╠Ū║¼┴┐£yČ©▓╔ė├▌ņ═¬▒╚╔½Ę©[9]ĪŻ┐╔╚▄ąįĄ░░ū┘|║¼┴┐£yČ©▓╔ė├┐╝±R╦╣┴┴╦{▒╚╔½Ę©[9]ĪŻĖ¼░▒╦ß║¼┴┐£yČ©▓╔ė├╦ßąį▄ß╚²═¬▒╚╔½Ę©[9]ĪŻ▒¹Č■╚®Ż©MDAŻ®║¼┴┐£yČ©▓╔ė├┴“┤·░═▒╚═ū╦ßĘ©[9]ĪŻPOD╗Ņąį£yČ©▓╔ė├▒╚╔½Ę©[9]ĪŻ 2 ĮY╣¹┼cĘų╬÷ 2.1Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“┐╔╚▄ąį╠Ū║¼┴┐Ą─ė░Ēæ ┐╔╚▄ąį╠Ū║¼┴┐┼cų▓╬’┐╣║«┴”į┌ČÓöĄų▓╬’ųą▒Ē¼F│÷š²ŽÓĻP[10]ĪŻ┐╔╚▄ąį╠Ū┐╔ęįį÷╝ėįŁ╔·┘|ØŌČ╚, Å─Č°ŲĄĮ┐╣├ō╦«ū„ė├╝░£p╔┘╝Ü░¹ā╚ĮY▒∙ÖCĢ■Ż¼ęį┤╦▀mæ¬═ŌĮńŁhŠ│ūā╗»Ż¼£p▌pę“Ą═£žČ°ę²ŲĄ─ā÷║”ĪŻ Å─łD 1 ┐╔ęį┐┤│÷į┌Ą═£ž├{Ų╚Ž┬,- 7Īµ╠Ä└Ē 24 h ┼c72h║¾┐╔╚▄ąį╠Ū║¼┴┐┬įėąį÷╝ė, ┼cī”šš▓Ņ«É▓╗┤¾ĪŻ▀@╩Ūė╔ė┌ų▓ųĻį┌5ĪµŽ┬Ģr¾wā╚ęčŠ█╝»ŽÓ«ööĄ┴┐Ą─╠ŪĘųĄųė∙Ą═£žĪŻį┌-18Īµ║═-25Īµ╠Ä└Ē║¾┐╔╚▄ąį╠Ū║¼┴┐Ž┬ĮĄŻ¼┐╔ęŖĖą╩▄Ą═£žā÷║”║¾Ż¼ų▓╬’┐╔╚▄ąį╠Ū║¼┴┐ėąŽ┬ĮĄĄ─┌ģä▌ĪŻ ├ń─ŠŠW,www.cqhuayin.com 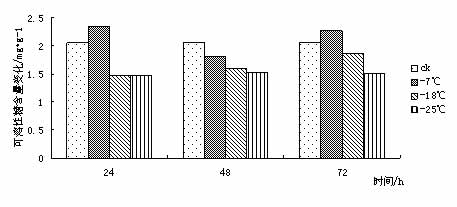 łD1 Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“┐╔╚▄ąį╠Ū║¼┴┐Ą─ė░Ēæ 2.2Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“┐╔╚▄ąįĄ░░ū┘|║¼┴┐Ą─ė░Ēæ ų▓╬’į┌─µŠ│Ž┬╩¦╦«é¹║”╩Ū╩ūꬥ─[10]ĪŻ┐╔╚▄ąįĄ░░ū┘|Ą─į÷╝ė┐╔ęį╩°┐`Ė³ČÓĄ─╦«Ęų,═¼Ģr┐╔ęį£p╔┘įŁ╔·┘|ā╚ĮY▒∙Č°é¹║”ų┬╦└Ą─ÖCĢ■ĪŻ Å─łD2┐╔ęį┐┤│÷į┌Ą═£ž├{Ų╚║¾Ż¼¢|Š®ę░▄į└“ėū├ńĒgŲż▓┐ųą┐╔╚▄ąįĄ░░ū│╩Ž╚╔Ž╔²║¾Ž┬ĮĄĄ─┌ģä▌Ż¼į┌-18Īµ╠Ä└Ē24hĪó72ąĪĢr║¾┐╔╚▄ąįĄ░░ū┘|║¼┴┐▀_ĄĮūŅ┤¾ųĄĪŻ┐╔ęŖ┐╔╚▄ąįĄ░░ū║¼┴┐Ą─į÷╝ė╩Ū¢|Š®ę░▄į└“┐╣├ō╦«Īó£p╔┘įŁ╔·┘|ā╚ĮY▒∙Ą─ųžę¬╩ųČ╬ų«ę╗ĪŻ 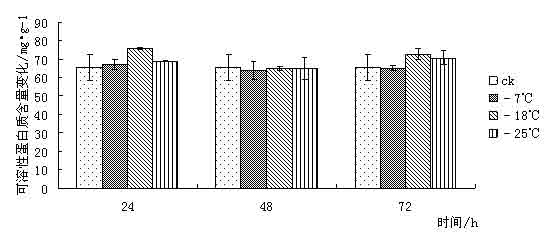 łDČ■ Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“┐╔╚▄ąįĄ░░ū┘|║¼┴┐Ą─ė░Ēæ 2.3Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“POD╗ŅąįĄ─ė░Ēæ š²│ŻŪķørŽ┬Ż¼ų▓╬’¾wā╚┤µį┌ų°ėąą¦Ą─▒Żūo├ĖŽĄĮy(PODĄ╚)Ż¼ęįŪÕ│²╗Ņąį觥╚ėąČŠ╬’┘|ĪŻ«öų▓╬’¾wė÷ĄĮĄ═£ž├{Ų╚ĢrŻ¼▀@ĘNŪÕ│²ÖCųŲŠ═Ģ■╩▄ūĶŻ¼ī¦ų┬¾wā╚╗Ņąį觥─Ęe└█Ż¼įņ│╔ī”─żĄ─é¹║”║═ī”┤¾ĘųūėĄ─ŲŲē─ĪŻ▀MČ°ī¦ų┬┤·ųx╣”─▄Ą─═╦╗»║═╝Ü░¹Ą─╦└═÷ĪŻ Å─łD3┐╔ęį┐┤│÷Ż¼į┌Ą═£ž├{Ų╚║¾Ż¼¢|Š®ę░▄į└“ėū├ńĒgŲż▓┐POD╗ŅąįŠ∙ėą╦∙į÷╝ėĪŻį┌-25╠Ä└Ē║¾Ą─24hŻ¼48hŻ¼72hĘųäe▀_ĄĮūŅ┤¾ųĄŻ¼▒Ē¼F│÷¢|Š®ę░▄į└“ī”Ą═£žėąų°▌^ÅŖĄ─▀mæ¬─▄┴”ĪŻ 999ųąć°├ń─ŠŠW,999miaomu.com 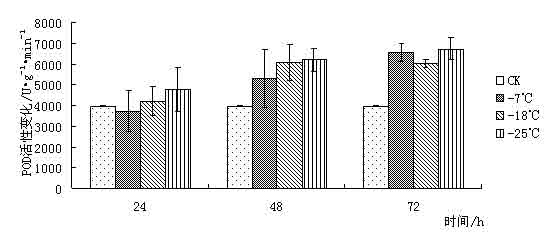 łD3 Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“POD╗ŅąįĄ─ė░Ēæ 2.4Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“ė╬ļxĖ¼░▒╦ß║¼┴┐Ą─ė░Ēæ Ė¼░▒╦ß╩Ūų▓╬’¾wā╚ę╗ĘNųžę¬Ą─ØB═Ėš{╣Ø╬’┘|,╦³Ą─Ęe└█ī”▀mæ¬─µŠ│ėąųžę¬ęŌ┴x,┐╔ęįĘ┤ė│ų▓╬’Ą─╩▄Ų╚║”ĀŅørĪŻę“┤╦,Ųõ║¼┴┐ūā╗»┐╔ęįū„×ķų▓╬’ī”─µŠ│├{Ų╚Ą─ę╗ĘN╔·└Ē╔·╗»ųĖś╦[11]ĪŻ ╚ńłD4╦∙╩ŠŻ¼Ą═£ž├{Ų╚Ž┬Ż¼¢|Š®ę░▄į└“ėū├ńĒgŲż▓┐ė╬ļxĖ¼░▒╦ß║¼┴┐├„’@į÷╝ėĪŻļSų°£žČ╚Ą─▓╗öÓĮĄĄ═, ė╬ļxĖ¼░▒╦ß║¼┴┐ėąųØuį÷╝ė║¾ĮĄĄ═Ą─┌ģä▌▓óį┌-18Īµ╠Ä└Ē48║¾▀_ĄĮūŅ┤¾ųĄĪŻ┐╔ęŖ¢|Š®ę░▄į└“═©▀^į÷╝ėė╬ļxĖ¼░▒╦ßĮ^ī”║¼┴┐üĒ╠ßĖ▀Ųõ┐╣║«ąįĪŻ 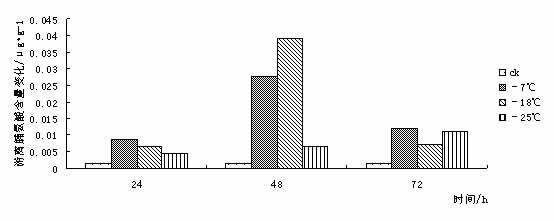 łD4Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“ė╬ļxĖ¼░▒╦ß║¼┴┐Ą─ė░Ēæ 2.5Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“MDA║¼┴┐Ą─ė░Ēæ ▒¹Č■╚® (MDA)╩Ū╔·╬’─żŽĄĮy─żų¼▀^č§╗»«a╬’ų«ę╗,Ųõ║¼┴┐▓╗āH▒Ē╩Š▀^č§╗»│╠Č╚║═─żŽĄĮyĄ─é¹║”│╠Č╚,Č°Ūęę▓┐╔ęįķgĮėĄž▒Ē╩ŠĮM┐Śųąūįė╔╗∙Ą─║¼┴┐[ 12 ]ĪŻ Å─łD5┐╔ęį┐┤│÷Ż¼į┌24h╠Ä└ĒŽ┬Ż¼ļSų°£žČ╚Ą─Ž┬ĮĄMDA║¼┴┐│÷¼FŽ╚╔²Ė▀║¾ĮĄĄ═Ą─┌ģä▌Ż¼šf├„Ą═£ž├{Ų╚Ž┬,ų¼┘|░l╔·▀^č§╗»ū„ė├,╝Ü░¹─ż╩▄ōpĪŻį┌48hŻ¼72h╠Ä└ĒŽ┬Ż¼Ą═£ž├{Ų╚║¾Ą─¢|Š®ę░▄į└“ėū├ńĒgŲż▓┐MDA║¼┴┐Š∙ėą╦∙Ž┬ĮĄŻ¼▀@šf├„ļSų°├{Ų╚ĢrķgĄ─š²│ŻŻ¼¢|Š®ę░▄į└“ųØuą▐Å═╝Ü░¹─żŻ¼ęį▀_ĄĮ┐╣║«Ą──┐Ą─ĪŻ ųąć°├ń─ŠŠW,999miaomu.com 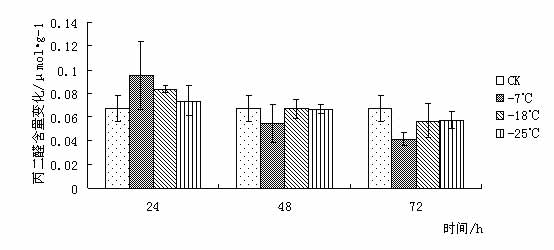 łD5Ą═£ž├{Ų╚ī”¢|Š®ę░▄į└“MDA║¼┴┐Ą─ė░Ēæ 3 ėæšō Ą═£ž▒∙ā÷Ģ■ę²Ųę╗ŽĄ┴ą╝Ü░¹┤·ųx╣”─▄Ą─Ė─ūā,╚╦éā▓╔ė├┐╔╚▄ąį╠Ū║¼┴┐Īó┐╔╚▄ąįĄ░░ū┘|║¼┴┐ĪóĖ¼░▒╦ß║¼┴┐ĪóSOD╗ŅąįĪóPOD╗ŅąįĪó▒¹Č■╚®║¼┴┐ĪóNR╗ŅąįĄ╚蹊┐┴ų─Š┐╣║«ąį[13]ĪŻ 3.1┐╔╚▄ąį╠Ū║¼┴┐┼c¢|Š®ę░▄į└“Ą─┐╣║«ąį 蹊┐ĮY╣¹▒Ē├„┐╔╚▄ąį╠Ū┐╔ęįį÷╝ėįŁ╔·┘|ØŌČ╚, Å─Č°ŲĄĮ┐╣├ō╦«ū„ė├╝░£p╔┘╝Ü░¹ā╚ĮY▒∙ÖCĢ■,ī”įŁ╔·┘|,ā÷├¶ĖąĄ░░ū┘|Īó┼╝┬ōę“ūėĄ╚ŲĄĮ▒Żūoū„ė├[10]ĪŻį┌░l╔·Ą═£žĢrŻ¼┤¾▓┐Ęųų▓╬’Č╝Ģ■Ęe└█ę╗Č©┴┐Ą─┐╔╚▄ąį╠ŪüĒ▀mæ¬Ą═£žÄ¦üĒĄ──µŠ│ĪŻ▒ŠīŹ“×ĮY╣¹▒Ē├„į┌-7ĪµĄ═£ž╠Ä└ĒŽ┬¢|Š®ę░▄į└“┐╔╚▄ąį╠Ū║¼┴┐ėą╦∙į÷╝ėŻ¼šf├„į┌-7ĪµĢr¢|Š®ę░▄į└“┐╔═©▀^╔²Ė▀Ųõ┐╔╚▄ąį╠Ū║¼┴┐üĒį÷╝ėįŁ╔·┘|ØŌČ╚Å─Č°Ąųė∙Ą═£ž├{Ų╚ĪŻ 3.2┐╔╚▄ąįĄ░░ū┘|║¼┴┐┼c¢|Š®ę░▄į└“Ą─┐╣║«ąį 蹊┐ĮY╣¹▒Ē├„┐╔╚▄ąįĄ░░ū┘|Ą─į÷╝ė┐╔ęį╩°┐`Ė³ČÓĄ─╦«Ęų,═¼Ģr┐╔ęį£p╔┘įŁ╔·┘|ā╚ĮY▒∙Č°é¹║”ų┬╦└Ą─ÖCĢ■[10]ĪŻ▒Šįć“×ĮY╣¹▒Ē├„į┌Ą═£ž├{Ų╚ĢrŻ¼¢|Š®ę░▄į└“═©▀^┐╔╚▄ąįĄ░░ūĄ─Ęe└█Ż¼Å─Č°ĮĄĄ═╦«ä▌Ż¼į÷ÅŖØB═Ėš{┐ž─▄┴”ĪŻ 3.3 POD╗Ņąį┼c¢|Š®ę░▄į└“Ą─┐╣║«ąį š²│ŻŪķørŽ┬Ż¼ų▓╬’¾wā╚┤µį┌ų°ėąą¦Ą─▒Żūo├ĖŽĄĮyŻ¼ęįŪÕ│²╗Ņąį觥╚ėąČŠ╬’┘|ĪŻ«öų▓╬’¾wė÷ĄĮĄ═£ž├{Ų╚ĢrŻ¼▀@ĘNŪÕ│²ÖCųŲŠ═Ģ■╩▄ĄĮūĶĄKŻ¼ī¦ų┬¾wā╚╗Ņąį觥─Ęe└█Ż¼įņ│╔ī”─żĄ─é¹║”║═┤¾ĘųūėĄ─ŲŲē─Ż¼╩╣DNA«a╔·ōpé¹Ż¼ė░ĒæĄ░░ū┘|Ą─║Ž│╔┼cĘĆČ©Ż¼▀MČ°įņ│╔┤·ųx╣”─▄Ž¹╩¦Ģr║═╝Ü░¹╦└═÷Ż¼ę╗░ŃšJ×ķŻ¼Ą═£ž├{Ų╚Ž┬POD╗Ņąį▒Ē¼F×ķŽ╚╔²║¾ĮĄ[14]. ųąć°├ń─ŠŠW,999miaomu.com ▒Šįć“×ųąŻ¼į┌Ą═£ž├{Ų╚Ž┬¢|Š®ę░▄į└“POD╗Ņąį│╩Ž╚╔Ž╔²║¾Ž┬ĮĄ┌ģä▌Ż¼ŪęļSų°ĢrķgĄ─į÷ķLėą▌^┤¾Ę∙Č╚Ą─╔²Ė▀Ż¼▒Ē├„¢|Š®ę░▄į└“į┌Ą═£ž├{Ų╚Ž┬▒Żūo├Ė┐╔ęįŠS│ų▌^Ė▀╦«ŲĮŻ¼Å─Č°▒▄├Ō┴╦╗Ņąį觥╚ūįė╔╗∙Ą─┤¾┴┐Ęe└█Ż¼£p▌p┴╦ų▓╬’¾wā╚╗Ņąįč§Ä¦üĒĄ─é¹║”ĪŻ 3.4ė╬ļxĖ¼░▒╦ß║¼┴┐┼c¢|Š®ę░▄į└“Ą─┐╣║«ąį 蹊┐ĮY╣¹▒Ē├„Ż¼Ė¼░▒╦ßį┌ų▓╬’┐╣ā÷ųąŠ▀ėąųžę¬ū„ė├,ļSų°£žČ╚Ą─▓╗öÓĮĄĄ═,ų▓╬’╚~Ų¼ųąĖ¼░▒╦ß║¼┴┐│╩├„’@į÷╝ė┌ģä▌ĪŻįĮ┐╣║«Ą─ŲĘĘNį÷╝ėĄ─▒ČöĄįĮĖ▀ĪŻ[11].Ė¼░▒╦ß╩Ūų▓╬’¾wā╚╦«╚▄ąįūŅ┤¾Ą─░▒╗∙╦ßų«ę╗Ż¼į┌Ą═£ž├{Ų╚Ž┬Ż¼ė╬ļxĖ¼░▒╦ߥ─Ęe└█į┌▀mæ¬─µŠ│ųąŲĄĮę╗Č©ū„ė├ĪŻ▒Šįć“×ųąŻ¼į┌Ą═£ž├{Ų╚Ž┬ė╬ļxĖ¼░▒╦ß║¼┴┐│╩Ž╚╔Ž╔²║¾Ž┬ĮĄ┌ģä▌Ż¼į┌-18Īµ╠Ä└Ē24hĄ─ĮMĘųųą▀_ĄĮūŅ┤¾ųĄŻ¼šf├„ė╬ļxĖ¼░▒╦ߥ─į÷╝ėĢr¢|Š®ę░▄į└“┐╣║«Ę┤æ¬Ą─ę╗ĒŚųžę¬╩ųČ╬ĪŻ 3.5 MDA║¼┴┐┼c¢|Š®ę░▄į└“Ą─┐╣║«ąį ▒¹Č■╚® (MDA)╩Ū╔·╬’─żŽĄĮy─żų¼▀^č§╗»«a╬’ų«ę╗,Ųõ║¼┴┐▓╗āH▒Ē╩Š▀^č§╗»│╠Č╚║═─żŽĄĮyĄ─é¹║”│╠Č╚,Č°Ūęę▓┐╔ęįķgĮėĄž▒Ē╩ŠĮM┐Śųąūįė╔╗∙Ą─║¼┴┐ĪŻäó┐ĪėóĄ╚į┌蹊┐Ą═£ž├{Ų╚ī”č®╦╔─żų¼▀^č§╗»╬’╝░▒Żūo├ĖĄ─ė░ĒæĢr,ūCīŹ┴╦ų▓╬’¾wā╚ MDA║¼┴┐┐╔ęįĘ┤ė│ų▓╬’─żé¹║”╝░╦ź└Ž│╠Č╚[11].▒Šįć“×ųąŻ¼Ą═£ž├{Ų╚Ž┬MDA║¼┴┐ļSų°ĢrķgĄ─ūā╗»│╩¼FŽ╚╔²Ė▀║¾ĮĄĄ═Ą─┌ģä▌Ż¼▀@▒Ē├„į┌Ą═£žŽ┬¢|Š®ę░▄į└“╝Ü░¹─ż╩▄ĄĮę╗Č©│╠Č╚Ą─ŲŲē─Ż¼║¾į┌PODĄ╚├ĖĄ─ū„ė├Ž┬ųØu╩╣╗Ņąįč§┤·ųxĄ─ŲĮ║Ō╗ųÅ═ĪŻšf├„ÖC¾w┐╣č§╗»ŽĄĮyĄ─š{╣Ø─▄┴”▌^ÅŖĪŻ ųąć°├ń─ŠŠW,999miaomu.com į┌┐╔╚▄ąį╠ŪĪó┐╔╚▄ąįĄ░░ū║¼┴┐į÷╝ė║═Ė¼░▒╦ßĪóPODĄ╚▒Żūo├ĖĄ─╣▓═¼ū„ė├Ž┬,¢|ĘĮ¢|Š®ę░▄į└“Š═─▄ī”Ą═£ž├{Ų╚«a╔·▌^ÅŖĄ─Ąų┐╣─▄┴”, Ą½╬ęéā═¼ĢrąĶųĖ│÷╔Ž╩÷╦∙╩╣ė├Ą─ 5éĆųĖś╦ųą,ų╗ėąė╬ļxĖ¼░▒╦ߊ▀ėąśO’@ų°▓Ņ«É,Ųõ╦¹ 4éĆųĖś╦Š∙▓Ņ«É▓╗’@ų°ĪŻšf├„į┌įō蹊┐ųąė╬ļxĖ¼░▒╦ß╩Ū£yČ©ų▓╬’┐╣║«ąįĄ─ųžę¬ųĖś╦ĪŻæ¬įōųĖ│÷,Š▀ėą┐╣║«┴”Ą─ų▓╬’ę╗Č©┤µį┌ų°┐╣║«╗∙ę“[15], į┌Ą═£žŽ┬▒Ē▀_│÷╔·└Ē║═ą╬æBĮYśŗĄ─┐╣ąįĪŻ╔·└Ē┐╣ąįę▓▓╗╩Ūå╬ę╗ę“ūėĄ─ū„ė├,┐╔─▄╩ŪŠC║ŽĄ─╔·└Ēę“╦ž[16]ĪŻę“┤╦╔Ž╩÷£yįćĄ─ÄūĘN╔·└Ēę“ūė┼c¢|Š®ę░▄į└“┐╣║«ąįėąę╗Č©ĻPŽĄ,╩Ūʱ┼cŲõ╦³ĘĮ├µėąĻPŽĄ▀Ćėą┤²▀Mę╗▓Į╠ĮėæĪŻ į┌įō蹊┐ųą,ėąįSČÓ╔ą▓╗─▄┤_Č©Ą─šJūR,Ę┤æ¬┴╦ų▓╬’┐╣║«ąįÖCųŲĄ─Å═ļsąį,ąĶę¬▀Mąą╔Ņ╚ļ蹊┐ĪŻų▓╬’╔·╗Ņį┌╣ŌĪó£žĪó ╦«Īó ÜŌĪó ¤ß╣▓═¼ū„ė├Ą─ŁhŠ│ųą,Ą═£ž├{Ų╚ų╗╩ŪŲõųąčąŠ┐Ą─ę╗éĆĘĮ├µ,ę“┤╦į┌╝ėÅŖå╬ę╗ę“ūė蹊┐Ą─╗∙ĄA╔Ž,蹊┐ļpę“ūė╗“ČÓę“ūė╣▓═¼ū„ė├,▓┼─▄▀_ĄĮę²ĘN│╔╣”║═▀mĄž▀mśõĄ──┐ś╦ĪŻ ģó┐╝╬─½I [1] ųąć°┐ŲīWį║ų▓╬’蹊┐╦∙. ųąć°Ė▀Ą╚ų▓╬’łDĶbĄ┌╚²āį[M]. ▒▒Š®Ż║┐ŲīW│÷░µ╔ńŻ¼1985. 338. [2] ÓŹ╚fŌx. ųąć°śõ─ŠųŠĄ┌Č■ŠĒ[M]. ▒▒Š®Ż║ųąć°┴ųśI│÷░µ╔ńŻ¼1985. 12. 1 602ŻŁ1604. ųąć°├ń─ŠŠW,999miaomu.com [3] ĖĄ┴óć°Ż¼ĻÉ╠ČŪÕ.ųąć°Ė▀Ą╚ų▓╬’Ą┌┴∙ŠĒ[M]. ŪÓŹu│÷░µ╔ńŻ¼2003Ż¼10. 29. [4] ążÅ═├„Ż¼į°ųŠ╣ŌŻ¼ŚŅśÕŻ¼Ą╚. ¢|Š®ę░▄į└“ĘNūėė═ĀIB│╔Ęų蹊┐[J]. ╠ņ╚╗«a╬’蹊┐┼cķ_░lŻ¼2005Ż¼17Ż©3Ż®Ż║344ŻŁ345. [5] ģŪ┐╦▀xŻ¼į°ųŠ╣ŌŻ¼ų▄ąĪŲĮŻ¼Ą╚. ¢|Š®ę░▄į└“ę░═Ōš{▓ķł¾Ėµ[J]. ĮŁ╬„┴ųśI┐Ų╝╝Ż¼2002Ż©2Ż®Ż║25ŻŁ27. [6] ┤„Ģį²ł. ¢|Š®ę░▄į└“╝░Ę▒ė²╝╝ąg[J]. ╠žĘNĮøØ·äėų▓╬’Ż¼2002Ż©6Ż®Ż║21. [7]ć└ŪÓ,±Rė±Ø²,╩®Į©▄Ŗ,Ą╚.Ą═£ž├{Ų╚ī”3 ĘN─┴▓▌ėū├ń┐╣ąį╔·└ĒųĖś╦Ą─ė░Ēæ[J ].ŪÓ║Ż┤¾īWīWł¾,2007 ,25(1) :54-57. [8]└Ķ├„,└ŅĖŻąŃ,±R¤©│╔,Ą╚.ŽŃ─Š╔Åī”Č╠ĢrĄ═£ž├{Ų╚╠Ä└ĒĄ─╔·└Ē╔·æBĒææ¬[J ].▒▒ĘĮł@╦ć,2006(1) :37-39. [9]Åłų╬░▓ĪóĻÉš╣ėŅŻ¼ų▓╬’╔·└ĒīWīŹ“×╝╝ągŻ¼╝¬┴ų┤¾īW│÷░µ╔ńŻ¼2008.7 100Ż¼110Ż¼182Ż¼192. [10]äóūµņ„Ą╚.ų▓╬’┐╣ąį╔·└ĒīW.▒▒Š®:ųąć°▐rśI│÷░µ╔ń,199 [11]ķZųął@, ĮčąŃæ.▓╗═¼ŲĘĘNūŽ░▀─ĄĄżĄ─┐╣║«ąį蹊┐┼c▒╚▌^[J ].░▓╗š▐rśI┐ŲīW,2009, 37 (24) :11511 - 11513. [12]äó┐Īėó,ę”┐ŲįŲ,±Tę½’w,Ą╚.Ą═£ž├{Ų╚ī”č®╦╔─żų¼▀^č§╗»╝░▒Żūo├ĖĄ─ė░Ēæ[ J ] .╔Į╬„▐rśI┤¾īWīWł¾, 2004 (4) : 399 - 400 . ųąć°├ń─ŠŠW,999miaomu.com [13]═§└¹,žSš,Åł¢|īÄ.ć°ā╚┴ų─Š┐╣║«ąį蹊┐Ą─▀Mš╣╝░š╣═¹[J].╔Į¢|┴ųśI┐Ų╝╝,2002(1):48-49. [14] ūT┤¾║Ż,┴_ą┬┴x,╔│éź.Ą═£ž├{Ų╚Ž┬4éĆ▄┘▐Żę²▀MŲĘĘN SODĪó POD╗ŅąįĄ─ūā╗»[J ].ųąć°’Ģ┴Ž,2007(1) :25-31. [15]9. Nordin et. al. Separate signal pathways regulate the expression of a low-temperature- induced gene in Arabidopsis thaliana(L.) Heynh, Plant Molecular Biology, 1991,(16): 1061-1071 [16]╚fŪÕ┴ųĄ╚.é╚Į▒K╗©│╔ųĻ─Ļ╔·ķL╣Ø┬╔╔·└Ē╠žąįĄ─蹊┐.ų▓╬’蹊┐,1996,(3):351-355. 999├ń─ŠŠW,www.cqhuayin.com ū„š▀Ż║═§║Į ążč┼ ī¦Ä¤Ż║ė„ĘĮłA 999ųąć°├ń─ŠŠW,999miaomu.com |
| ╔Žę╗Ų¬Ż║╦»╔ÅĄ─įį┼Ó┼c╣▄└ĒłD Ž┬ę╗Ų¬Ż║┬Āšf▀^╠Ų▌┼Ųčå߯┐ |
CopyRight 2017 www.cqhuayin.com ╩±ICPéõ17029831╠¢-2ųąć°├ń─ŠŠW
All Rights Reserved.┐═Ę■Qó┘ó┌
ó█
ŠWšŠĄžłDRRSĄžłD░┘Č╚ĄžłD